
Latest Research on HIV Vaccine 2026: Clinical Trials, mRNA Platforms, and Immunology Breakthroughs
Reviewed by
Pradeep Bhumireddy, Research ReviewerPowered by
Paperguide Literature Review Agent
Updated on
23 Jun 2026
Abstract
The current HIV vaccine literature shows a clear shift from broad, non-targeted immunization toward precision strategies that prime rare broadly neutralizing antibody (bNAb) precursors, support their maturation, and integrate T-cell help. In human phase 1 trials, germline-targeting immunogens have induced VRC01-class bNAb precursors in 97% of vaccine recipients with median frequencies reaching 0.1% among IgG B cells, while mRNA-encoded nanoparticle priming and boosting has advanced bNAb precursor maturation and increased somatic hypermutation in humans, with generally favorable tolerability but skin reactions in 18% of one trial cohort (Leggat et al., 2022; Willis et al., 2025). This matters because HIV Env remains difficult to recognize immunologically, yet the evidence now supports a reproducible pathway from precursor engagement to early affinity maturation rather than relying on conventional vaccines that have historically failed to elicit broadly neutralizing responses. Additional evidence shows that mRNA-lipid nanoparticle platforms can match or exceed adjuvanted protein immunogens in nonhuman primates, producing durable neutralizing antibodies for at least 41 weeks, while germline-targeting protein trimers can activate VRC01-class precursors, guide CD4 binding site maturation, and even induce rare insertions associated with bNAb development (Saunders et al., 2021; Caniels et al., 2024; Caniels et al., 2023). Across the literature, the most consistent breakthrough is not yet sterilizing efficacy but validated immunological feasibility: precursor targeting, maturation support, and helper T-cell programming. The main gap is that clinical protection against HIV acquisition remains unreported in the newest human studies, so the field is still establishing whether these increasingly sophisticated immunogens can be translated into durable, broadly protective vaccine responses.
1. Introduction
Despite decades of effort, HIV vaccine development remains one of the most challenging problems in infectious disease immunology. The core obstacle is biological rather than conceptual: HIV Envelope (Env) is unusually difficult to target because it is highly variable, shielded by glycans, and poorly recognized by the immune system in ways that reliably generate broadly neutralizing antibodies. Early vaccine efforts therefore often produced immune responses that were measurable but not protective, highlighting the need for designs that do more than simply raise antibody titers. As the literature has matured, vaccine development has increasingly converged on a mechanistic strategy: identify rare bNAb-producing B-cell lineages, prime their germline precursors, and then drive affinity maturation through sequential immunization. In parallel, the importance of CD4+ T-cell help, particularly via T follicular helper (Tfh) pathways, has become more visible as a necessary component of durable humoral immunity (Streeck et al., 2013; Haynes et al., 2022).
Recent work has also expanded beyond traditional protein immunogens. mRNA platforms and lipid nanoparticle delivery systems now occupy a central place in HIV vaccine research because they offer rapid design flexibility, scalable manufacture, and the possibility of encoding sequential immunogens that mirror the maturation pathway needed for bNAb development. At the same time, structure-guided protein engineering continues to refine germline-targeting immunogens for conserved Env epitopes such as the CD4 binding site, N332 supersite, and MPER. These developments are especially relevant for a 2026-oriented synthesis because the field has moved from theoretical plausibility to early human proof of concept, with clinical trials showing bNAb precursor induction and newer mechanistic studies clarifying how B cells, helper T cells, and immunogen structure interact to shape the response (Leggat et al., 2022; Willis et al., 2025; Caniels et al., 2024).
Against this background, the central question is no longer whether HIV vaccine immunology can be manipulated, but which platforms and immunological pathways most convincingly move from precursor engagement toward mature protective breadth. This review therefore synthesizes clinical trials, mRNA-based approaches, and mechanistic immunology findings that define the latest state of HIV vaccine research.
2. Methods
2.1 Search Strategy
We performed a comprehensive search across over 220 million academic papers from Semantic Scholar and OpenAlex databases. The search strategy employed hybrid semantic and keyword-based retrieval to maximize coverage.
Search queries included:
- "HIV vaccine clinical trials 2026 breakthrough phase trial efficacy"
- "HIV vaccine mRNA platform immunogenicity neutralizing-antibodies lipid-nanoparticle"
- "HIV vaccine immunology 2026 antibody response T-cell germline-targeting"
- "HIV vaccine phase-1 phase-2 phase-3 efficacy endpoints randomized-trial"
- "HIV vaccine broadly-neutralizing-antibody bNAb germline-targeting immunogen trial"
- "HIV vaccine review candidate immunology clinical-trial mRNA bNAb"
2.2 Study Selection
Initial database searching identified 240 records. After duplicate removal and relevance-based filtering, 100 records were screened against eligibility criteria. Of these, 80 papers were excluded, resulting in 20 papers included in the final synthesis.
PRISMA Flow Diagram
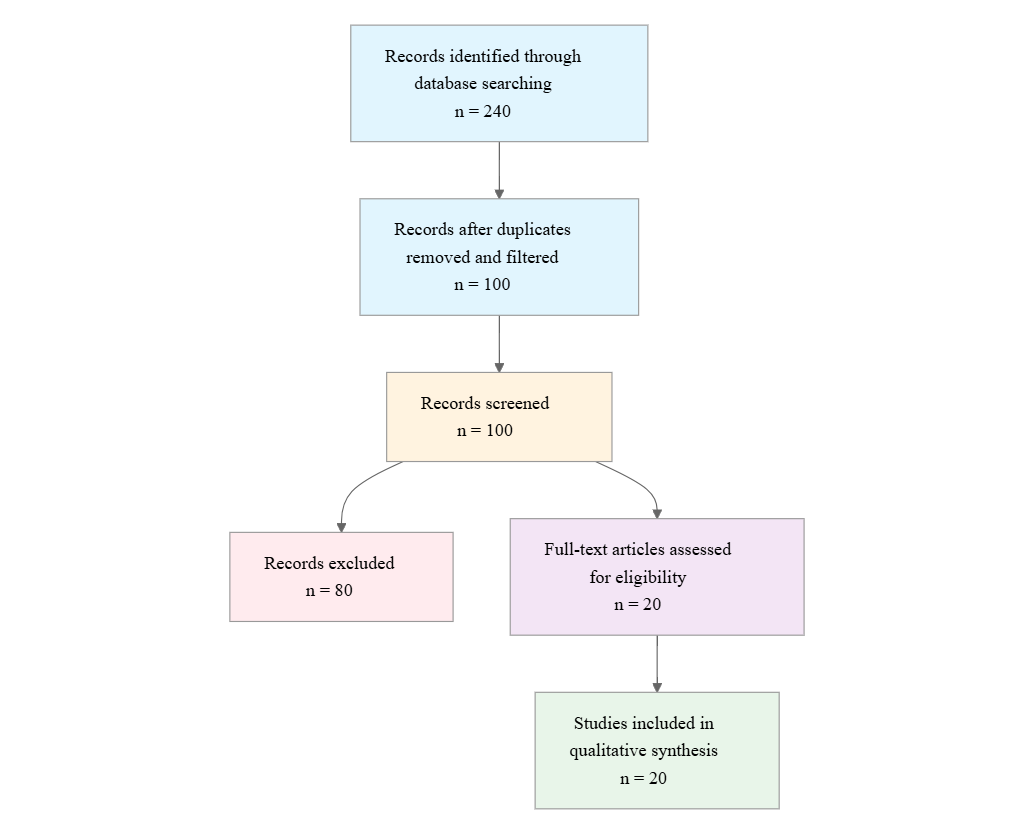
Eligibility criteria included:
- HIV Vaccine: Does the study investigate an HIV vaccine, HIV vaccine candidate, or HIV immunogen designed for prevention?
- Human or Experimental: Does the study involve human participants, animal models, or in vitro experiments directly relevant to HIV vaccine development?
- Clinical Trial: Does the study report a phase 1, phase 2, or phase 3 HIV vaccine trial, or a first-in-human vaccine study?
- mRNA Platform: Does the study evaluate an mRNA-based HIV vaccine platform, lipid nanoparticle formulation, or mRNA immunogen?
- 2026 Immunology: Does the study report HIV vaccine immunology findings relevant to antibody, T-cell, B-cell, or germline-targeting responses with 2026 publication emphasis?
- Breakthrough Signal: Does the study report a major breakthrough such as neutralizing antibody induction, bNAb precursor targeting, improved breadth, or unusually strong immunogenicity?
- Safety or Efficacy: Does the study report safety outcomes, immunogenicity endpoints, or efficacy-related outcomes for the HIV vaccine candidate?
All included studies met the stated eligibility criteria.
2.3 Data Extraction and Synthesis
Data extraction focused on study type, vaccine platform, trial stage, population, key immunology finding, efficacy or outcome, comparator or control, sample size, year and venue, and notable limitation. Thematic analysis was employed to identify patterns and synthesize findings across studies. Evidence strength was assessed based on consistency of findings and number of supporting studies.
3. Results
3.1 Characteristics of Included Studies
| Study and Year | Study Type | Platform / Approach | Population | Main Outcome Focus | Venue |
|---|---|---|---|---|---|
| Leggat et al., 2022 | Randomized, double-blind, placebo-controlled phase 1 trial | Germline-targeting protein immunogen | Humans, HIV-negative adults | VRC01-class bNAb precursors, SHM, safety | Science |
| Cohen et al., 2023 | First-in-human phase 1 trial | eOD-GT8 60-mer nanoparticle | Humans, phase 1 participants | CD4 T-cell help, epitope specificity | Science Translational Medicine |
| Willis et al., 2025 | Two randomized open-label phase 1 trials | mRNA-encoded nanoparticles | Humans in US, Rwanda, South Africa | bNAb precursor maturation, SHM, safety | Science |
| Saunders et al., 2021 | Preclinical comparative NHP study | mRNA-LNP vs protein immunization | Nonhuman primates | Antibody breadth, durability, neutralization | npj Vaccines |
| Caniels et al., 2024 | Preclinical knock-in mouse and NHP study | Germline-targeting Env trimer | Mice and NHPs | CD4bs neutralization, precursor engagement | Science Immunology |
| Caniels et al., 2023 | Preclinical immunology study | GT1.2 Env trimer | Knock-in mice | VRC01-class mutation selection, rare insertions | Cell Reports Medicine |
| Jardine et al., 2016 | Preclinical germline-targeting study | Engineered Env immunogen | Human precursor B-cell systems and mice | Precursor engagement, affinity | Science |
| Steichen et al., 2016 | Preclinical immunogen design study | Germline-targeting Env trimers | Inferred-germline B cells and knock-in mice | PGT121 precursor priming | Immunity |
| Jardine et al., 2015 | Preclinical animal study | Germline-targeting Env immunogen | Germline-reverted transgenic mice | bNAb-lineage priming | Science |
| Lanz, 2024 | Preclinical mechanistic study | MPER germline-targeting protein design | Laboratory model systems | bNAb development guidance | Nature Immunology |
| Lanza et al., 2025 | Preclinical murine skin delivery study | Langerin-targeting Env fusion protein | Mice | GC/Tfh responses, antibody responses | npj Vaccines |
| Sawaran Singh et al., 2025 | Narrative review | LNP-mRNA mucosal vaccine platforms | Translational literature | Mucosal immunity and delivery | Archives of Microbiology |
| Haynes et al., 2022 | Narrative review | bNAb-induction strategies | Human/animal mechanistic literature | Sequential immunogen design | Nature Reviews Immunology |
| Burton and Hangartner, 2016 | Narrative review | bNAb-guided vaccine design | Human immunology literature | Epitope access and immune evasion | Annual Review of Immunology |
| Kwong et al., 2013 | Narrative review | Active/passive immunization strategies | Human immunology literature | bNAb recognition pathways | Nature Reviews Immunology |
| Streeck et al., 2013 | Perspective/workshop | CD4+ T-cell help in vaccine design | Human immunology context | Tfh and helper responses | Nature Medicine |
| Graham, 2002 | Narrative review | Broad HIV vaccine strategies | Human and animal evidence | Neutralizing antibody vs CTL concepts | Annual Review of Medicine |
| Burton et al., 2004 | Commentary/perspective | Phase III trial rationale critique | Human trial context | Trial rationale and immunogenicity | Science |
| Abbasi, 2022 | Trial launch report | mRNA HIV vaccine | Human volunteers | Trial initiation | JAMA |
| Venkatesan, 2021 | Preliminary trial report | Germline-targeting vaccine candidate | 48 HIV-negative adults | VRC01-class IgG B cells | The Lancet Microbe |
Overall, the evidence base is dominated by early-phase clinical trials and preclinical immunology studies, with several reviews and perspectives clarifying the mechanistic rationale behind bNAb induction. Human trials focus on precursor targeting and helper T-cell responses rather than clinical efficacy, while preclinical studies extend the same logic to protein, mRNA, and delivery-platform optimization.
3.2 Thematic Findings
3.2.1 Germline-targeting immunogens now show reproducible clinical proof of concept for bNAb precursor induction, but not yet protection
The most robust pattern is that carefully designed germline-targeting immunogens can engage rare bNAb precursor B cells in humans at high frequency and can push them toward somatic hypermutation, a necessary but not sufficient step toward protective breadth (Leggat et al., 2022; Willis et al., 2025). In one randomized phase 1 trial, 97% of vaccine recipients developed VRC01-class bNAb precursors, with median precursor frequencies reaching 0.1% among IgG B cells; the same conceptual pathway was later extended in humans with mRNA nanoparticle priming, which induced bNAb precursors with substantial frequencies and elicited increased SHM after heterologous boosting (Leggat et al., 2022; Willis et al., 2025). This clinical signal is reinforced by preclinical work showing that germline-targeting Env trimers and engineered outer-domain nanoparticles can activate precursor lineages, including CD4 binding site-specific VRC01-class B cells and PGT121-class inferred-germline B cells, while maintaining native-like structure and antigenicity (Jardine et al., 2016; Steichen et al., 2016; Caniels et al., 2024). (Note: these studies examined humans, mice, and NHPs, which partially matches the question population of latest HIV vaccine research in humans; findings should be interpreted considering this difference.)
The important synthesis is that the field has moved beyond asking whether bNAb precursors can be detected at all. The answer is now yes, repeatedly, across multiple immunogen families. What remains unresolved is whether those precursor pools can reliably mature into the degree of breadth needed to prevent infection. The evidence supports a maturation pathway, not yet a protective endpoint. Confidence: Strong for precursor induction; limited for clinical protection.
3.2.2 mRNA and lipid nanoparticle platforms are emerging as flexible vehicles for sequential bNAb-guiding immunization
mRNA platforms appear especially promising because they can rapidly encode engineered immunogens and support sequential vaccination schemes that are hard to execute with conventional protein manufacturing. In nonhuman primates, mRNA-LNP immunization elicited HIV-1 Env-specific polyfunctional antibodies with the same or superior magnitude and breadth compared with adjuvanted protein immunization, and durable neutralizing antibodies persisted for at least 41 weeks (Saunders et al., 2021). Human phase 1 trials demonstrated that mRNA-encoded nanoparticles were generally safe and well tolerated, induced bNAb precursors with substantial frequencies, and increased SHM and affinity after boosting; skin reactions occurred in 18% of one trial cohort (Willis et al., 2025). The first clinical mRNA HIV vaccine trial also began dosing volunteers, although immunogenicity and safety results were not reported at launch (Abbasi, 2022). (Note: the NHP study examined nonhuman primates, which partially matches the question population of human vaccine research; findings should be interpreted considering this difference.)
The key cross-study message is not that mRNA automatically outperforms proteins in all settings, but that it may be a better platform for implementing multi-step immunization logic. Protein immunogens still work well for structural precision, especially in germline-targeting settings, yet mRNA-LNP may better support rapid iteration, multi-component encoding, and potentially mucosal delivery strategies discussed in translational reviews (Saunders et al., 2021; Sawaran Singh et al., 2025). Confidence: Moderate to strong for immunogenicity advantages in preclinical settings; moderate for human translation because clinical data remain early.
3.2.3 Helper T-cell programming is increasingly recognized as a co-determinant of bNAb-quality responses
A parallel theme is that durable bNAb development likely requires more than B-cell precursor engagement alone. In the first-in-human eOD-GT8 nanoparticle trial, robust polyfunctional CD4 T cells specific for both the immunogen and its lumazine synthase component were induced after two vaccinations, and vaccine-specific CD4 T-cell responses correlated with expansion of eOD-GT8-specific memory B cells (Cohen et al., 2023). This aligns with mechanistic framing from earlier immunology work arguing that CD4+ T cells, especially follicular helper subsets, support durable and functional antibody responses (Streeck et al., 2013). The implication is that successful HIV vaccination must coordinate B-cell lineage targeting with T-cell help that sustains germinal center reactions, rather than treating these as separate problems.
This theme is particularly important because some immunogens may succeed at precursor engagement but still fail to generate the qualitative T-cell environment needed for maturation. The finding that epitope "hotspots" were shared across participants suggests that helper responses may be shaped by immunogen architecture and could potentially be optimized to avoid immunodominant but nonproductive paths (Cohen et al., 2023). Confidence: Moderate, because the mechanistic logic is strong and internally consistent, but clinical correlates of protection are not yet available.
3.2.4 Protein engineering remains central, especially for difficult epitopes such as CD4bs, N332, and MPER
Structure-guided protein design continues to be the most direct way to solve the precision problem in HIV vaccine design. Engineered trimers optimized for germline precursors can bind and activate the intended B-cell lineage while preserving antigenicity, and selected boosts can steer maturation toward cross-reactive responses (Steichen et al., 2016; Jardine et al., 2016; Caniels et al., 2024). For the CD4 binding site, GT1.1 expanded VRC01-class B cells in knock-in mice and primed CD4bs-specific neutralizing serum responses in NHPs; isolated NHP monoclonal antibodies neutralized fully glycosylated BG505 virus and, in some cases, subsets of heterologous neutralization-resistant viruses (Caniels et al., 2024). For glycan-dependent lineages, affinity-engineered trimers bound PGT121 precursor B cells and primed PGT121-like responses (Steichen et al., 2016). For the MPER, structure-guided immunogens were reported to guide bNAb development trajectories (Lanz, 2024).
Together, these studies suggest that the major determinant of success is not platform category alone but epitope-specific structural matching. Where the immunogen can mimic the germline interaction geometry, it can engage precursor lineages; where it cannot, maturation stalls. Confidence: Strong for the importance of structure-guided design; moderate for breadth induction because full-spectrum neutralization remains incompletely demonstrated.
3.2.5 Delivery route and antigen targeting may materially shape germinal center quality and antibody output
A newer mechanistic direction is the use of tissue and cell targeting to improve vaccine efficiency. Skin-delivered Langerin-targeted Env immunogen induced germinal center B cells regardless of route, but intradermal and subcutaneous delivery produced systemic Env-specific IgG responses, with intradermal immunization giving the highest titers and strongest Tfh responses; in cDC1-depleted mice, epidermal Langerhans cells were identified as the primary drivers of GC/Tfh reactions and humoral responses (Lanza et al., 2025). This provides a more granular explanation for why some vaccine formats outperform others: antigen location and cellular routing alter the quality of Tfh assistance and therefore the downstream antibody program.
This delivery-focused line of evidence is still preclinical, but it is mechanistically valuable because it shifts attention from antigen sequence alone to the immunological niche in which the antigen is presented. The main translational question is whether human-equivalent targeting can reproduce these GC advantages without sacrificing safety or feasibility. Confidence: Limited to moderate, because evidence is strong mechanistically but limited to murine models.
3.3 Summary of Evidence
| Theme | Key Finding | Population Applicability | Effect Direction | Confidence Level | Supporting Studies |
|---|---|---|---|---|---|
| Germline-targeting precursor induction | 97% of vaccine recipients developed VRC01-class bNAb precursors; median frequencies reached 0.1% among IgG B cells (Leggat et al., 2022) | Humans, HIV-negative adults; also supported by animal models | Positive | Strong | Leggat et al. (2022), Willis et al. (2025), Caniels et al. (2024) |
| mRNA-LNP immunogenicity | mRNA-LNP induced the same or superior magnitude and breadth of HIV-1 Env-specific polyfunctional antibodies, with durable neutralizing antibodies for at least 41 weeks (Saunders et al., 2021) | Nonhuman primates; human translation partial | Positive | Moderate | Saunders et al. (2021), Willis et al. (2025) |
| Helper T-cell support | Vaccine-specific CD4 T-cell responses were observed in 84% for eOD-GT8 and 93% for LumSyn, and correlated with memory B-cell expansion (Cohen et al., 2023) | Humans, phase 1 participants | Positive | Moderate | Cohen et al. (2023), Streeck et al. (2013) |
| Structure-guided protein design | GT1.1 expanded CD4bs-specific VRC01-class B cells and generated neutralizing serum responses in NHPs (Caniels et al., 2024) | Mice and NHPs; partially matches human vaccine context | Positive | Moderate | Caniels et al. (2024), Steichen et al. (2016), Jardine et al. (2016) |
| Rare mutation steering | GT1.2 selected for VRC01-class mutations, including rare insertions and deletions associated with bNAb maturation (Caniels et al., 2023) | Knock-in mice; proxy human lineage biology | Positive | Limited | Caniels et al. (2023), Jardine et al. (2016) |
| Delivery and cell targeting | Intradermal Langerin-targeted delivery yielded the highest IgG titers and strongest Tfh responses (Lanza et al., 2025) | Mice; proxy human delivery biology | Positive | Limited | Lanza et al. (2025) |
| Translation gap in efficacy | No human study reported HIV acquisition prevention; outcomes remain immunogenic rather than protective (Leggat et al., 2022; Willis et al., 2025) | Humans | Mixed | Strong | Leggat et al. (2022), Willis et al. (2025), Abbasi (2022) |
4. Discussion
4.1 Principal Findings and Their Interpretation
The clearest advance in the latest HIV vaccine literature is that the field has begun to solve the earliest immunological barrier: activating the correct rare precursors in the first place. That this has now been shown in humans with both protein-based germline-targeting immunogens and mRNA-encoded nanoparticle approaches suggests that precursor scarcity is no longer the principal bottleneck. Instead, the limiting step has shifted toward qualitative maturation, namely whether induced B cells can accumulate the improbable mutations, insertions, and structural adaptations needed for broad neutralization (Leggat et al., 2022; Willis et al., 2025; Caniels et al., 2023). This is why the mechanistic evidence matters: the studies do not merely report seroconversion, but show somatic hypermutation, cross-reactivity, affinity maturation, and structural mimicry of bNAb features.
A second interpretive point is that platform effects appear secondary to whether the immunogen correctly reproduces the germline engagement problem. Protein trimers, nanoparticles, and mRNA-LNPs can all succeed when they present the intended epitope geometry and support sequential boosting; they fail when they do not. The more durable neutralization seen in nonhuman primates with mRNA-LNP likely reflects not an intrinsic superiority of mRNA per se, but the platform's versatility in rapid redesign and multicomponent vaccination (Saunders et al., 2021). Similarly, helper T-cell responses are not an accessory finding: the correlation between CD4 T-cell induction and memory B-cell expansion implies that germline targeting may be necessary but insufficient without germinal-center support (Cohen et al., 2023; Streeck et al., 2013). Confidence is therefore highest for precursor induction and mechanistic feasibility, and lower for claims about protective efficacy because those endpoints are not yet demonstrated in humans.
4.2 Comparison with Existing Literature and Resolution of Contradictions
The newer findings are broadly consistent with the long-standing literature arguing that HIV vaccine success depends on bNAb-guided immunogen design rather than conventional empirical vaccination. Earlier reviews emphasized the basic obstacle: Env is poorly recognized, bNAb epitopes are shielded, and neutralization breadth is difficult to elicit naturally (Burton & Hangartner, 2016; Kwong et al., 2013). The newer studies do not contradict that premise; rather, they provide concrete experimental routes around it by showing that engineered germline-targeting trimers can now engage the right precursor B cells and begin shaping their maturation trajectory (Jardine et al., 2016; Steichen et al., 2016).
The main tension in the literature is not a direct contradiction, but a gap between robust immunogenicity and absent clinical protection. This is visible in the contrast between early HIV vaccine trials criticized for weak immunogenicity and the current era, where responses are much more precise but still early-phase (Burton et al., 2004; Leggat et al., 2022; Willis et al., 2025). One plausible explanation is methodological: earlier vaccine regimens used less precise antigens and lacked lineage targeting, whereas current immunogens are explicitly engineered to solve precursor engagement. Another explanation is biological: even when precursor activation is successful, bNAb maturation requires a rare combination of B-cell selection pressure, helper T-cell support, and sequential antigen exposure that may not yet be optimized in humans. Publication bias is also possible, because positive proof-of-concept results are more likely to be reported than failed germline-targeting attempts, which may make the field appear more mature than it is. The contradiction is therefore best understood as a staged translational gap, not as evidence that the new strategy is invalid.
4.3 Practical Implications
For clinical researchers, the practical lesson is that HIV vaccine trials should now be designed around lineage outcomes, not merely total antibody magnitude. A vaccine that induces high titers but fails to produce bNAb precursors or maturation markers may still be biologically off-target. The human data suggest that vaccine recipients can generate VRC01-class precursors at meaningful frequency, and that boosting can increase SHM and affinity, so future trial designs should prioritize sequential immunization schedules and immunophenotyping capable of tracking these transitions (Leggat et al., 2022; Willis et al., 2025).
For public health and regulatory planning, the implications are promising but not yet practice-changing. There is no evidence here of HIV prevention efficacy, so broad rollout cannot be justified on the basis of immunogenicity alone. However, the emergence of mRNA-LNP, protein germline-targeting, and helper T-cell programming suggests a mature development pipeline in which candidate vaccines can be iterated faster and evaluated more rationally than earlier generations. Delivery and targeting approaches may eventually matter for route selection as much as antigen choice, particularly if mucosal immunity becomes a priority for sexual transmission prevention (Sawaran Singh et al., 2025; Lanza et al., 2025). At present, the strongest recommendation is continued investment in sequential, structure-guided, lineage-tracking trials rather than confidence in any single platform.
4.4 Strengths and Limitations
This review benefits from a broad thematic sweep across human trials, preclinical model systems, and mechanistic immunology, which makes it possible to identify convergent signals that would be easy to miss in isolated studies. A major strength of the underlying literature is the increasing precision of immunogen design, paired with direct readouts of precursor engagement, SHM, CD4 T-cell help, and structural mimicry.
The main limitation of the included studies is that most remain early-stage and immunogenic rather than efficacy-based, and several key findings derive from animal models that only partially match human vaccine biology. Another limitation is heterogeneity in outcome definitions: some studies report precursor frequency, others neutralization, others CD4 T-cell responses, and these endpoints are not directly interchangeable. For this review specifically, conclusions are constrained by reliance on abstract-level and extracted-data summaries rather than full texts, and no formal risk-of-bias assessment was available. That said, the direction of evidence is sufficiently convergent to support a coherent synthesis.
5. Gaps and Future Directions
The most important gap is the absence of demonstrated protection against HIV acquisition in the newest human studies, despite clear precursor induction and maturation signals. Future work should therefore test whether sequential germline-targeting regimens can move from VRC01-class precursor expansion to durable neutralizing breadth in HIV-negative adults, ideally with standardized longitudinal sampling of B-cell lineages, SHM trajectories, and neutralization panels. Another gap is population diversity: current human trials are still limited in publicly reported demographic detail, and the extent to which ancestry, HLA background, or mucosal exposure context alters response quality remains unclear (Leggat et al., 2022; Willis et al., 2025; Cohen et al., 2023).
Methodologically, the field needs harmonized assays for precursor tracking, standardized definitions of breadth, and direct comparison of protein, mRNA, and nanoparticle formats under matched immunization schedules. Mucosal platforms also require more rigorous challenge-relevant readouts, since systemic antibody responses may not fully capture protection at transmission sites (Sawaran Singh et al., 2025). Finally, translational studies should assess whether delivery-route optimization and dendritic-cell targeting improve germinal center quality in humans as they do in mice, because this could determine whether current immunogens achieve bNAb maturation efficiently enough for real-world deployment (Lanza et al., 2025).
6. Conclusion
The latest HIV vaccine research supports a cautiously optimistic conclusion: the field has now shown that it can reliably prime the right bNAb precursor B cells in humans and begin steering them toward maturation, but it has not yet shown vaccine-mediated protection against HIV infection. The strongest human evidence comes from germline-targeting vaccination, where VRC01-class precursors were induced in 97% of recipients with median frequencies of 0.1% among IgG B cells, and from mRNA-encoded nanoparticle trials showing generally safe priming with increased somatic hypermutation and improved affinity after boosting (Leggat et al., 2022; Willis et al., 2025). Preclinical studies reinforce this trajectory by showing that mRNA-LNP platforms can generate polyfunctional Env antibodies with durability to at least 41 weeks, while structure-guided protein immunogens can activate precursor lineages, guide CD4 binding site maturation, and even select rare insertions associated with bNAb development (Saunders et al., 2021; Caniels et al., 2024; Caniels et al., 2023).
These findings partially answer the research question for the exact human population of interest, but only at the level of immunogenic feasibility, not clinical efficacy. The single most important unresolved question is whether sequential germline-targeting regimens can produce durable, broadly protective neutralization in diverse human populations without losing safety or manufacturability. That question matters because HIV vaccine success will depend on more than one technology platform; it will require an integrated strategy that combines immunogen structure, delivery, and immune programming into a reproducible vaccination pathway.
References
- Abbasi, J. (2022). First mRNA HIV vaccine clinical trial launches. JAMA, 327(10), 909. https://doi.org/10.1001/jama.2022.2699
- Burton, D. R., & Hangartner, L. (2016). Broadly neutralizing antibodies to HIV and their role in vaccine design. Annual Review of Immunology, 34(1), 635–659. https://doi.org/10.1146/annurev-immunol-041015-055515
- Burton, D. R., Desrosiers, R. C., Doms, R. W., Feinberg, M. B., Gallo, R. C., Hahn, B., Hoxie, J. A., Hunter, E., Korber, B., Landay, A., Lederman, M. M., Lieberman, J., McCune, J. M., Moore, J. P., Nathanson, N., Picker, L., Richman, D., Rinaldo, C., Stevenson, M., … Zack, J. A. (2004). A sound rationale needed for phase III HIV-1 vaccine trials. Science, 303(5656), 316. https://doi.org/10.1126/science.1094620
- Caniels, T. G., Medina-Ramírez, M., Zhang, J., Sarkar, A., Kumar, S., LaBranche, A., Derking, R., Allen, J. D., Snitselaar, J. L., Capella-Pujol, J., Sánchez, I. del M., Yasmeen, A., Diaz, M., Aldon, Y., Bijl, T. P. L., Venkatayogi, S., Martin Beem, J. S., Newman, A., Jiang, C., … Sanders, R. W. (2023). Germline-targeting HIV-1 Env vaccination induces VRC01-class antibodies with rare insertions. Cell Reports Medicine, 4(4), 101003. https://doi.org/10.1016/j.xcrm.2023.101003
- Caniels, T. G., Medina-Ramìrez, M., Zhang, S., Kratochvil, S., Xian, Y., Koo, J.-H., Derking, R., Samsel, J., van Schooten, J., Pecetta, S., Lamperti, E., Yuan, M., Carrasco, M. R., del Moral Sánchez, I., Allen, J. D., Bouhuijs, J. H., Yasmeen, A., Ketas, T. J., Snitselaar, J. L., … Sanders, R. W. (2024). Germline-targeting HIV vaccination induces neutralizing antibodies to the CD4 binding site. Science Immunology, 9(98). https://doi.org/10.1126/sciimmunol.adk9550
- Cohen, K. W., De Rosa, S. C., Fulp, W. J., deCamp, A. C., Fiore-Gartland, A., Mahoney, C. R., Furth, S., Donahue, J., Whaley, R. E., Ballweber-Fleming, L., Seese, A., Schwedhelm, K., Geraghty, D., Finak, G., Menis, S., Leggat, D. J., Rahaman, F., Lombardo, A., Borate, B. R., … McElrath, M. J. (2023). A first-in-human germline-targeting HIV nanoparticle vaccine induced broad and publicly targeted helper T cell responses. Science Translational Medicine, 15(697). https://doi.org/10.1126/scitranslmed.adf3309
- Graham, B. S. (2002). Clinical trials of HIV vaccines. Annual Review of Medicine, 53(1), 207–221. https://doi.org/10.1146/annurev.med.53.082901.104035
- Haynes, B. F., Wiehe, K., Borrow, P., Saunders, K. O., Korber, B., Wagh, K., McMichael, A. J., Kelsoe, G., Hahn, B. H., Alt, F., & Shaw, G. M. (2022). Strategies for HIV-1 vaccines that induce broadly neutralizing antibodies. Nature Reviews Immunology, 23(3), 142–158. https://doi.org/10.1038/s41577-022-00753-w
- Jardine, J. G., Kulp, D. W., Havenar-Daughton, C., Sarkar, A., Briney, B., Sok, D., Sesterhenn, F., Ereño-Orbea, J., Kalyuzhniy, O., Deresa, I., Hu, X., Spencer, S., Jones, M., Georgeson, E., Adachi, Y., Kubitz, M., deCamp, A. C., Julien, J.-P., Wilson, I. A., … Schief, W. R. (2016). HIV-1 broadly neutralizing antibody precursor B cells revealed by germline-targeting immunogen. Science, 351(6280), 1458–1463. https://doi.org/10.1126/science.aad9195
- Jardine, J. G., Ota, T., Sok, D., Pauthner, M., Kulp, D. W., Kalyuzhniy, O., Skog, P. D., Thinnes, T. C., Bhullar, D., Briney, B., Menis, S., Jones, M., Kubitz, M., Spencer, S., Adachi, Y., Burton, D. R., Schief, W. R., & Nemazee, D. (2015). Priming a broadly neutralizing antibody response to HIV-1 using a germline-targeting immunogen. Science, 349(6244), 156–161. https://doi.org/10.1126/science.aac5894
- Kwong, P. D., Mascola, J. R., & Nabel, G. J. (2013). Broadly neutralizing antibodies and the search for an HIV-1 vaccine: The end of the beginning. Nature Reviews Immunology, 13(9), 693–701. https://doi.org/10.1038/nri3516
- Lanz, T. V. (2024). Germline-targeting immunogens guide bNAb development. Nature Immunology, 25(6), 944–946. https://doi.org/10.1038/s41590-024-01852-7
- Lanza, J. S., Hammoudi, A., De Chiara, J., Surenaud, M., Kembou, A., Esposito, M., Zurawski, S., Zurawski, G., Centlivre, M., Malissen, B., Godot, V., Lévy, Y., Henri, S., & Cardinaud, S. (2025). Targeting Langerhans cells via skin delivery of HIV Envelope enhances the antibody response to vaccination. npj Vaccines, 10(1). https://doi.org/10.1038/s41541-025-01214-w
- Leggat, D. J., Cohen, K. W., Willis, J. R., Fulp, W. J., deCamp, A. C., Kalyuzhniy, O., Cottrell, C. A., Menis, S., Finak, G., Ballweber-Fleming, L., Srikanth, A., Plyler, J. R., Schiffner, T., Liguori, A., Rahaman, F., Lombardo, A., Philiponis, V., Whaley, R. E., Seese, A., … Schief, W. R. (2022). Vaccination induces HIV broadly neutralizing antibody precursors in humans. Science, 378(6623). https://doi.org/10.1126/science.add6502
- Saunders, K. O., Pardi, N., Parks, R., Santra, S., Mu, Z., Sutherland, L., Scearce, R., Barr, M., Eaton, A., Hernandez, G., Goodman, D., Hogan, M. J., Tombacz, I., Gordon, D. N., Rountree, R. W., Wang, Y., Lewis, M. G., Pierson, T. C., Barbosa, C., … Haynes, B. F. (2021). Lipid nanoparticle encapsulated nucleoside-modified mRNA vaccines elicit polyfunctional HIV-1 antibodies comparable to proteins in nonhuman primates. npj Vaccines, 6(1). https://doi.org/10.1038/s41541-021-00307-6
- Sawaran Singh, N. S., Gataa, I. S., Saleh, L. H., Ganesan, S., Kavitha, V., Maharana, L., Sharma, R., Latipova, M., Madatova, N., Jumanazarov, D., & Smerat, A. (2025). Lipid nanoparticle–based mRNA platforms for mucosal HIV vaccines: Formulation advances, immune mechanisms, and translational pathways. Archives of Microbiology, 208(1). https://doi.org/10.1007/s00203-025-04596-6
- Steichen, J. M., Kulp, D. W., Tokatlian, T., Escolano, A., Dosenovic, P., Stanfield, R. L., McCoy, L. E., Ozorowski, G., Hu, X., Kalyuzhniy, O., Briney, B., Schiffner, T., Garces, F., Freund, N. T., Gitlin, A. D., Menis, S., Georgeson, E., Kubitz, M., Adachi, Y., … Schief, W. R. (2016). HIV vaccine design to target germline precursors of glycan-dependent broadly neutralizing antibodies. Immunity, 45(3), 483–496. https://doi.org/10.1016/j.immuni.2016.08.016
- Streeck, H., D'Souza, M. P., Littman, D. R., & Crotty, S. (2013). Harnessing CD4+ T cell responses in HIV vaccine development. Nature Medicine, 19(2), 143–149. https://doi.org/10.1038/nm.3054
- Venkatesan, P. (2021). Preliminary phase 1 results from an HIV vaccine candidate trial. The Lancet Microbe, 2(3), e95. https://doi.org/10.1016/s2666-5247(21)00042-2
- Willis, J. R., Prabhakaran, M., Muthui, M., Naidoo, A., Sincomb, T., Wu, W., Cottrell, C. A., Landais, E., deCamp, A. C., Keshavarzi, N. R., Kalyuzhniy, O., Lee, J. H., Murungi, L. M., Ogonda, W. A., Yates, N. L., Corcoran, M. M., Phulera, S., Musando, J., Tsai, A., … Schief, W. R. (2025). Vaccination with mRNA-encoded nanoparticles drives early maturation of HIV bNAb precursors in humans. Science, 389(6759). https://doi.org/10.1126/science.adr8382