
Latest Research on Type 1 Diabetes (2026): Causes, Insulin Delivery, Microneedles, and Immunotherapy
Reviewed by
Pradeep Bhumireddy, Research ReviewerPowered by
Paperguide Literature Review Agent
Updated on
23 Jun 2026
Abstract
Type 1 diabetes (T1D) is driven primarily by immune-mediated beta-cell destruction, with environmental triggers and genetic susceptibility shaping disease initiation, while therapeutic progress is strongest for insulin delivery innovation and immune intervention. Across the evidence base, insulin replacement remains foundational but incomplete, whereas teplizumab has shown the most clinically meaningful disease-modifying effect by delaying progression to clinical diabetes in high-risk individuals and preserving C-peptide in new-onset disease (Herold et al., 2019; Herold et al., 2013). Microneedle platforms have also advanced rapidly, including hollow, dissolving, coated, stimuli-responsive, and phase-transition systems that improve painless transdermal insulin delivery, accelerate release, and in some cases prolong normoglycemic time or approach hypodermic bioavailability (Gupta et al., 2009; Lee et al., 2016; Zhang et al., 2018; Y. Zhang et al., 2018; He et al., 2024; Yang et al., 2015; Wang et al., 2020; Chen et al., 2018). Mechanistic reviews converge on T cells as central effectors of beta-cell loss, but also emphasize that beta-cells, peri-islet Schwann cells, HLA-linked susceptibility, and environmental exposures such as viruses, diet, and microbiota contribute to pathogenesis (Burrack et al., 2017; Toren et al., 2021; Winer et al., 2003; Knip & Simell, 2012; Houeiss et al., 2022; Du et al., 2025; Carlsson, 2019). Collectively, the literature supports a dual trajectory for T1D care: immune modification to slow or prevent disease, and increasingly sophisticated insulin-delivery technologies to reduce treatment burden and improve glycemic control. However, direct causal evidence for environmental triggers remains limited, and most microneedle studies remain preclinical, limiting immediate translation.
1. Introduction
Type 1 diabetes is a lifelong autoimmune disease in which endogenous insulin production declines as pancreatic beta cells are progressively destroyed. Although exogenous insulin has transformed survival and metabolic control, it does not replace immune tolerance or fully prevent long-term complications, leaving a persistent need for therapies that address both the cause of disease and the burden of treatment (Chatenoud, 2010). Contemporary work shows that T1D is not a single-process disorder: autoreactive CD4+ and CD8+ T lymphocytes, environmental exposures, genetic susceptibility, and potentially beta-cell intrinsic vulnerability all shape the tempo of disease (Burrack et al., 2017; Knip & Simell, 2012; Toren et al., 2021; Du et al., 2025).
This complexity has driven two major research directions. One seeks to interrupt autoimmunity before irreversible beta-cell loss occurs, exemplified by anti-CD3 therapy with teplizumab, which delays progression in high-risk individuals and preserves residual beta-cell function after diagnosis (Herold et al., 2019; Herold et al., 2013). The other seeks to improve how insulin is delivered once the disease is established, especially through minimally invasive and smart transdermal systems. In this area, microneedle technologies have evolved from proof-of-concept hollow devices to dissolving, coated, phase-transition, and glucose-responsive patches designed to improve patient comfort, drug loading, release kinetics, and usability (Gupta et al., 2009; Lee et al., 2016; Zhang et al., 2018; Y. Zhang et al., 2018; He et al., 2024; Yang et al., 2015; Wang et al., 2020; Chen et al., 2018).
At the same time, mechanistic reviews increasingly argue that beta-cell destruction reflects more than passive immune attack. Environmental trigger hypotheses implicate enteroviruses, dietary antigens, microbiota shifts, inflammatory metabolic signatures, and endogenous retroviruses, while human and animal studies also suggest that peri-islet tissues and beta-cell heterogeneity may actively participate in pathogenesis (Knip & Simell, 2012; Houeiss et al., 2022; Winer et al., 2003; Toren et al., 2021; Du et al., 2025). Against this backdrop, the central question is how the newest evidence redefines the causes of T1D and the most promising treatment strategies spanning insulin, microneedles, and immunotherapy.
2. Methods
2.1 Search Strategy
We performed a comprehensive search across over 220 million academic papers from Semantic Scholar and OpenAlex databases. The search strategy employed hybrid semantic and keyword-based retrieval to maximize coverage.
Search queries included:
- "type-1-diabetes etiology pathogenesis autoimmune triggers beta-cell destruction genetics environmental"
- "type-1-diabetes insulin therapy basal bolus glycemic-control continuous glucose-monitoring HbA1c"
- "type-1-diabetes microneedle insulin delivery transdermal patch smart-release bioengineering"
- "type-1-diabetes immunotherapy prevention remission anti-CD3 teplizumab immune-modulation"
- "type-1-diabetes 2026 latest research review treatment causes clinical-trial advances"
- "type-1-diabetes systematic-review emerging therapies immunotherapy insulin delivery pathogenesis"
2.2 Study Selection
Initial database searching identified 240 records. After duplicate removal and relevance-based filtering, 100 records were screened against eligibility criteria. Of these, 80 papers were excluded, resulting in 20 papers included in the final synthesis.
PRISMA Flow Diagram
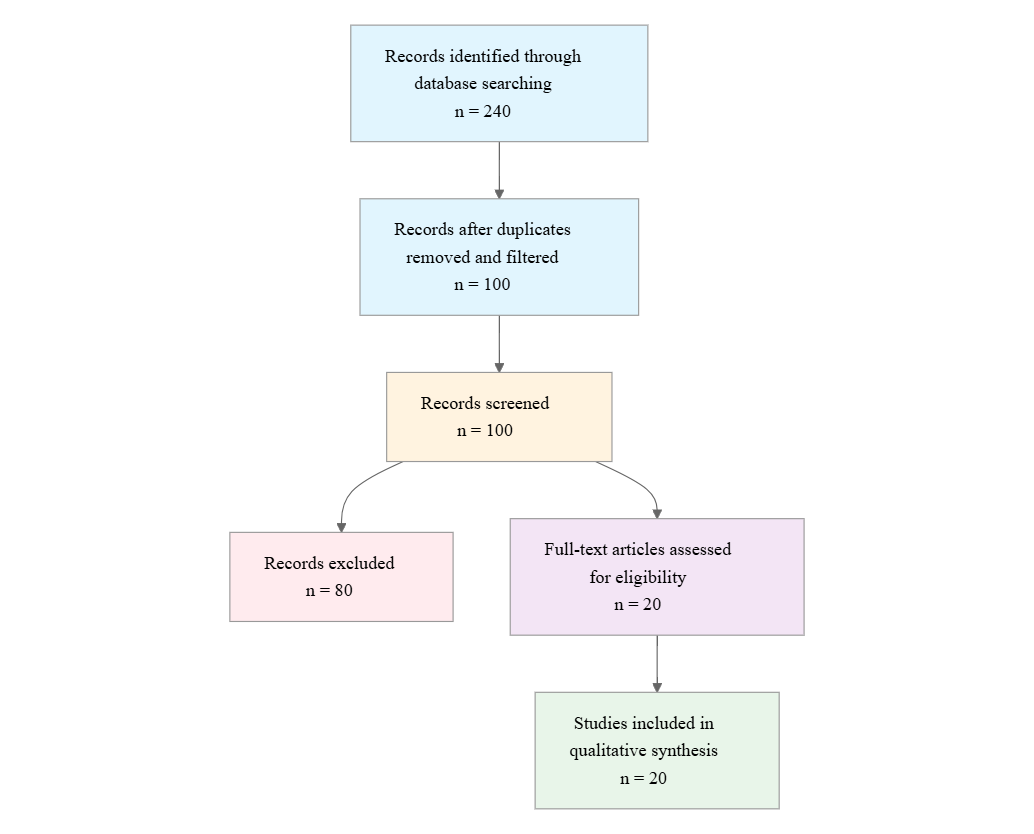
Eligibility criteria included:
- T1D Focus: Does the study focus on type 1 diabetes or clearly include a T1D-specific subgroup or T1D model?
- Human or Model: Does the study involve human participants, animal models, or in vitro systems relevant to T1D mechanisms or treatment?
- Causes or Treatment: Does the study address at least one of the following: causes/pathogenesis, insulin therapy, microneedle delivery, or immunotherapy for T1D?
- Recent Evidence: Does the study report research published in 2021-2026 or provide recent evidence relevant to current T1D research?
- Insulin Data: Does the study provide insulin-related findings such as dosing, delivery, glycemic control, or formulation performance?
- Microneedle Data: Does the study evaluate microneedle or transdermal delivery technology for insulin or other T1D-related agents?
- Immunotherapy Data: Does the study evaluate immunotherapy or immune modulation aimed at preventing, delaying, or treating T1D?
- Mechanism Data: Does the study report mechanistic findings about autoimmunity, beta-cell destruction, genetics, or environmental triggers in T1D?
All included studies met the stated eligibility criteria.
2.3 Data Extraction and Synthesis
Data extraction focused on the following variables:
- Research focus
- Study type
- Population/model
- Cause findings
- Insulin findings
- Microneedle findings
- Immunotherapy findings
- Key outcome
Thematic analysis was employed to identify patterns and synthesize findings across studies. Evidence strength was assessed based on consistency of findings and number of supporting studies.
3. Results
3.1 Characteristics of Included Studies
| Study and Year | Study Type | Population/Model | Key Focus | Primary Outcome/Endpoint |
|---|---|---|---|---|
| Winer et al. 2003 | Preclinical mechanistic investigation | NOD mice; NOD/SCID recipients; human prediabetes context referenced | Autoimmune destruction beyond beta cells | pSC-directed autoreactivity precedes beta-cell death; GFAP/S100β immunotherapy prevented diabetes |
| Knip & Simell 2012 | Narrative review | Human and mechanistic evidence synthesis | Environmental triggers | Enterovirus, dietary antigen, inflammation, microbiota hypotheses |
| Chatenoud 2010 | Narrative review | Review of T1D pathogenesis and immunotherapy | Anti-CD3 immune therapy | Anti-CD3 highlighted as distinctive immune-modulating strategy |
| Herold et al. 2013 | Randomized controlled trial | 52 humans with new-onset T1D | Teplizumab | Reduced 2-year C-peptide decline |
| Yang et al. 2015 | Preclinical device study | Engineered insulin-loaded patch platform | Phase-transition microneedles | Rapid dermal release; >20% bioavailability in pig models |
| Lee et al. 2016 | Preclinical device study | Diabetic mice; ex vivo skin testing | Two-layer dissolving microneedles | 50% release at 1 h; 80% by 5 h; RBA 85.7% |
| Zhang et al. 2017 | Preclinical device study | Diabetic SD rats | Alginate/maltose microneedles | Relative bioavailability 93.7±4.7% |
| Burrack et al. 2017 | Narrative review | Human and translational immunology evidence | T-cell-mediated beta-cell destruction | Autoimmunity and alloimmunity in T1D and transplantation |
| Chen et al. 2018 | Preclinical device study | Diabetic mice; cadaveric skin | Dissolving insulin patches | Rapid release and feasibility for clinical skin sites |
| Carlsson 2019 | Narrative review | Human adult LADA comparisons | Etiology overlap with T1D | HLA overlap and autoimmune pathogenesis in LADA |
| Herold et al. 2019 | Randomized clinical trial | Human relatives at risk for T1D | Teplizumab prevention/delay | Delayed progression to clinical T1D |
| Zhang et al. 2020 | Preclinical device study | Not specified | Glucose-responsive dissolving patch | Glucose-triggered insulin release |
| Wang et al. 2020 | Preclinical device study | Not specified | Coated microneedle patch | Rapid, painless insulin delivery |
| Zhang et al. 2021 | Preclinical device study | FITC-insulin skin-release testing | Dissolving polymer microneedles | ~90% tip localization; release within 2 min |
| Toren et al. 2021 | Narrative review | Human and experimental evidence synthesis | Beta-cell vulnerability and heterogeneity | Beta-cells actively contribute to disease progression |
| Houeiss et al. 2022 | Narrative review | Human and animal evidence synthesis | Environmental triggering | Viral, dietary, microbiota links to autoimmunity |
| He et al. 2024 | Preclinical device study | Diabetic mice / glucose-responsive testing | Triple-structure microneedle patch | Up to 24 h hypoglycemic effect; visible release completion |
| Du et al. 2025 | Review | Human and mechanistic evidence synthesis | Pathogenesis and treatment strategies | Genetics, environment, ERVs, and treatment overview |
| Foster et al. 2025 | Review | Clinical trial landscape synthesis | Emerging immunotherapies | Teplizumab as FDA-approved disease-modifying therapy |
| She et al. 2025 | Review | Clinical trial landscape synthesis | Emerging immunotherapies | Past 40 years of immunotherapy trials and future directions |
Overall, the literature splits into three dominant clusters: mechanistic work on immune and environmental causes, translational immunotherapy aimed at delaying beta-cell loss, and device engineering studies focused on transdermal insulin delivery. Human evidence is strongest for teplizumab and weakest for causal claims about environmental triggers, which remain largely inferential. Most microneedle studies are preclinical, but their designs increasingly address practical limitations such as incomplete release, poor loading, and pain.
3.2 Thematic Findings
3.2.1 T1D pathogenesis is consistently framed as immune-mediated, but the target tissue is broader and more dynamic than beta cells alone
Across mechanistic syntheses, T1D is presented as a progressive autoimmune disease driven by T-cell-mediated destruction of insulin-producing tissue (Burrack et al., 2017; Toren et al., 2021; Du et al., 2025). However, the evidence also broadens the disease concept beyond a simple beta-cell-only model. One mechanistic study showed that peri-islet Schwann cells are targeted early: pSC-autoreactive T- and B-cell responses appeared in 3- to 4-week-old NOD mice before detectable beta-cell death, and GFAP-specific T-cell lines transferred pathogenic peri-insulitis to NOD/SCID mice; immunotherapy with GFAP or S100β prevented diabetes (Winer et al., 2003). This suggests that the initiating autoimmune lesion may be peri-islet and neuroglial as well as endocrine. Parallel review evidence argues that beta-cells are not passive victims: beta-cell vulnerability, heterogeneity, and transcriptional responses may actively shape disease progression (Toren et al., 2021). In adult-onset autoimmune diabetes, LADA shares HLA-region genetic risk with T1D while also displaying insulin-resistance features, reinforcing that autoimmune beta-cell injury can coexist with metabolic modifiers (Carlsson, 2019). Confidence: Strong for immune-mediated destruction; moderate for the broader claim that non-beta-cell islet tissues and beta-cell-intrinsic states materially influence progression.
3.2.2 Environmental triggers remain plausible but unproven causal initiators, with viruses, diet, microbiota, and inflammatory profiles recurring across reviews
Environmental literature converges on a multi-hit model in which genetic susceptibility is necessary but insufficient. Reviews repeatedly identify enteroviruses as the leading infectious hypothesis, while also raising dietary antigens, microbiota shifts, and proinflammatory metabolic profiles as potential initiators or accelerators of autoimmunity (Knip & Simell, 2012; Houeiss et al., 2022; Du et al., 2025). The 2025 synthesis adds endogenous retroviruses and epigenetic modification to this framework (Du et al., 2025). Importantly, the evidence cited here is largely associative or animal-based rather than causal. Human observations link viral exposure, dietary habits, and microbiota changes to islet autoantibodies or overt diabetes, but prediction still relies mostly on autoantibody detection rather than validated exposure-response pathways (Houeiss et al., 2022). The strongest interpretation supported by the data is therefore not that any single exposure causes T1D, but that several environmental contexts may accelerate autoimmune initiation in susceptible hosts. Confidence: Moderate for a general environmental contribution; limited for any individual trigger as a proven causal factor.
3.2.3 Teplizumab is the clearest disease-modifying intervention, but benefit is partial and phenotype-dependent
Immune intervention evidence is far more mature than environmental causation evidence. In high-risk relatives, teplizumab delayed progression to clinical T1D (Herold et al., 2019), and in new-onset T1D it reduced 2-year C-peptide decline to −0.28 nmol/L [95% CI −0.36 to −0.20] versus −0.46 nmol/L [95% CI not fully shown] in controls, a 75% improvement (Herold et al., 2013). Reviews of anti-CD3 therapy emphasize why this matters: T1D is driven by autoreactive CD4+ and CD8+ T cells, so a targeted immune modulator is better aligned with pathogenesis than broad immunosuppression (Chatenoud, 2010; Foster et al., 2025). Yet the benefit is incomplete, as responses vary and duration is limited. The trial also noted that metabolic and immunologic baseline features distinguished responders from nonresponders, implying that disease stage and immune state likely mediate efficacy (Herold et al., 2013). Confidence: Strong that anti-CD3 therapy can delay disease and preserve beta-cell function; moderate that baseline phenotype can be used to predict who benefits most.
3.2.4 Microneedle insulin delivery is rapidly advancing toward painless, rapid, and smarter transdermal administration, but the evidence remains predominantly preclinical
The device literature shows remarkable convergence on one goal: making insulin delivery less painful and more physiologic without sacrificing dosing reliability. Hollow microneedles provided the first proof that bolus insulin can be delivered effectively to people with T1D in a minimally invasive manner (Gupta et al., 2009). Dissolving systems then improved localization and release control: gelatin/CMC patches released 50% of insulin by 1 h and 80% by 5 h, with relative bioavailability of 85.7% and pharmacologic availability of 95.6% in diabetic mice (Lee et al., 2016); alginate/maltose composite microneedles in diabetic SD rats achieved relative bioavailability of 93.7±4.7% and relative pharmacological availability of 94.1±5.6% (Zhang et al., 2018). Later systems pushed toward smarter control: glucose-responsive microneedles improved normoglycemic duration and achieved an approximately 0.3 cm² patch effect for up to 24 h in diabetic mice, while a triple-structure patch provided visible indication of release completion (He et al., 2024). Phase-transition patches exceeded 20% transdermal bioavailability in pig models and were engineered for complete removal without tip deposition (Yang et al., 2015). These effects are directionally consistent, but comparability is limited because models, endpoints, and formulations differ. Confidence: Moderate for feasibility and therapeutic promise; limited for comparative superiority among device classes.
3.2.5 Across microneedle platforms, the main engineering bottlenecks are drug loading, release completeness, and usability rather than raw skin penetration alone
A recurring theme is that successful microneedle translation depends less on whether the skin can be punctured and more on whether the device can store, release, and signal insulin delivery reliably. Conventional polymeric microneedles are repeatedly described as constrained by limited drug loading and incomplete release; newer designs address this with tip localization, glucose-responsive shells, inner propelling cores, and structural optimization (Y. Zhang et al., 2018; He et al., 2024; Yang et al., 2015). The dissolving PVA/sucrose system localized about 90% of FITC-insulin in conical tips and released it into skin within 2 min (Y. Zhang et al., 2018), whereas the glucose-responsive system explicitly targeted closed-loop-like release guidance via fading inner cores (He et al., 2024). This evolution implies that patient adherence and dose confidence are emerging as design priorities alongside pharmacokinetic performance. Confidence: Strong that engineering priorities are shifting toward complete, visible, and controllable delivery; moderate that these features will translate to clinical benefit.
3.3 Summary of Evidence
| Theme | Key Finding | Population Applicability | Effect Direction | Confidence Level | Supporting Studies |
|---|---|---|---|---|---|
| Immune-mediated pathogenesis | pSC autoreactivity preceded beta-cell death; GFAP/S100β immunotherapy prevented diabetes in NOD models | Mostly animal and translational immunology; partially matches human T1D | Negative for disease progression when targeted | Strong | Winer et al. (2003), Burrack et al. (2017), Toren et al. (2021) |
| Environmental triggering | Viral, dietary, microbiota, and inflammatory hypotheses recur, but causality remains unproven | Human observational and animal evidence; partially matches T1D population | Mixed | Moderate | Knip & Simell (2012), Houeiss et al. (2022), Du et al. (2025) |
| Teplizumab disease modification | C-peptide decline at 2 years was −0.28 nmol/L [95% CI −0.36 to −0.20] vs −0.46 nmol/L in controls | Human high-risk relatives and new-onset T1D; strong match | Positive | Strong | Herold et al. (2019), Herold et al. (2013), Foster et al. (2025) |
| Microneedle feasibility | Relative bioavailability reached 85.7% in mice, 93.7±4.7% in rats, and >20% in pigs depending on design | Mostly animal and proof-of-concept human study; partially matches T1D care context | Positive | Moderate | Gupta et al. (2009), Lee et al. (2016), Zhang et al. (2018) |
| Smart transdermal delivery | Glucose-responsive and visualized patches prolonged normoglycemic time and enabled release guidance | Mostly preclinical; partially matches T1D treatment population | Positive | Moderate | Y. Zhang et al. (2018), He et al. (2024), Wang et al. (2020) |
| Beta-cell and islet complexity | Beta-cells are active participants, and LADA shares HLA risk with T1D | Human and translational reviews; partially matches T1D population | Mixed | Moderate | Toren et al. (2021), Carlsson (2019), Burrack et al. (2017) |
4. Discussion
4.1 Principal Findings and Their Interpretation
The synthesis supports a clear hierarchy of evidence in T1D research. The most mature clinical advance is immune intervention, particularly teplizumab, which delays progression to clinical disease and slows loss of endogenous insulin secretion (Herold et al., 2019; Herold et al., 2013). This is biologically coherent because the central pathogenic process in T1D is autoimmune T-cell activity directed at pancreatic tissue (Burrack et al., 2017; Chatenoud, 2010). The fact that a targeted anti-CD3 strategy works where broad insulin replacement cannot modify disease course suggests that the intervention aligns with the disease driver rather than the downstream metabolic consequence. The responder subgroup findings further imply that immune therapy is most effective before beta-cell reserve is fully exhausted and when the inflammatory state is still modifiable (Herold et al., 2013).
The mechanistic literature extends this interpretation by showing that the autoimmune target may be broader than canonical beta-cell destruction alone. The early peri-islet Schwann cell findings in NOD mice indicate that islet neuroglial structures may participate in the initiation of local inflammation (Winer et al., 2003), while beta-cell heterogeneity may determine susceptibility to immune attack and progression rate (Toren et al., 2021). That combination helps explain why some disease-modifying strategies succeed only partially: they may dampen effector immunity but not fully reset the tissue-level susceptibility landscape. Environmental trigger hypotheses are best interpreted as upstream modifiers of this same immune circuit, not as stand-alone causes (Knip & Simell, 2012; Houeiss et al., 2022; Du et al., 2025).
By contrast, microneedle research addresses the clinical burden after diagnosis rather than the pathogenesis itself. The strongest synthesis finding here is engineering convergence around faster, less painful, and more reliable insulin delivery. Even though most evidence is preclinical, the progression from hollow to dissolving, glucose-responsive, and visually guided systems suggests that the field is solving practical barriers one by one: pain, incomplete release, and dose uncertainty (Gupta et al., 2009; Lee et al., 2016; He et al., 2024). The evidence is therefore strongest for feasibility and weakest for direct clinical superiority.
4.2 Comparison with Existing Literature and Resolution of Contradictions
The pattern of findings is broadly consistent with the established view of T1D as an autoimmune disease, but this synthesis adds two important refinements. First, the literature no longer treats beta cells as passive targets. The peri-islet Schwann cell data and beta-cell heterogeneity reviews suggest that tissue context helps initiate or amplify autoimmunity, which means that immune therapies aimed only at circulating lymphocytes may be necessary but insufficient (Winer et al., 2003; Toren et al., 2021). Second, the environmental literature has shifted from single-cause models toward a multi-exposure framework involving infections, diet, microbiota, and inflammation (Knip & Simell, 2012; Houeiss et al., 2022; Du et al., 2025). This is meaningful because it supports a multi-stage disease model in which genetic risk creates susceptibility, environmental exposures trigger immune activation, and local tissue responses shape progression.
The main contradiction is between strong mechanistic plausibility and weak causal proof for environmental triggers. Reviews repeatedly identify enteroviruses, dietary antigens, and microbiota changes, but they also acknowledge that prediction remains mostly based on autoantibodies rather than causal exposure evidence (Knip & Simell, 2012; Houeiss et al., 2022). This likely reflects heterogeneous exposure timing, imperfect exposure measurement, and the difficulty of observing preclinical autoimmunity in humans. It may also reflect publication bias, because positive associations with biologically plausible triggers are more likely to be emphasized than null findings. The evidence base therefore supports caution: these exposures are credible candidates, but not yet proven causes.
A second tension concerns microneedle translation. Device studies consistently show promising delivery metrics, yet the models are heterogeneous and often preclinical (Yang et al., 2015; Lee et al., 2016; Zhang et al., 2018; He et al., 2024). Apparent superiority across platforms may reflect differences in formulation, animal physiology, and endpoint selection rather than intrinsic device quality. As a result, the field has demonstrated feasibility more convincingly than it has demonstrated a clinically decisive winner.
4.3 Practical Implications
For clinicians, the most actionable implication is that T1D is increasingly stratifiable. Individuals at high risk, especially relatives with presymptomatic autoimmunity, may benefit from early immune intervention because teplizumab can delay clinical onset and preserve beta-cell function (Herold et al., 2019; Herold et al., 2013). For patients with established disease, the practical promise lies in smarter insulin delivery rather than cure: microneedle systems may reduce pain, simplify administration, and potentially improve adherence, especially if glucose-responsive or visually guided devices can be validated in humans (He et al., 2024; Wang et al., 2020).
From a public health perspective, the environmental literature argues for continued surveillance of viral and other upstream exposures in genetically susceptible groups, but it does not yet justify single-cause prevention policies (Knip & Simell, 2012; Houeiss et al., 2022; Du et al., 2025). The evidence supports exposure reduction and early detection strategies rather than precise trigger-based prevention. For regulators and device developers, the clearest message is that pain reduction alone is not enough; devices must also demonstrate reliable loading, complete release, stable insulin activity, and clinically meaningful glycemic control across realistic use conditions (Y. Zhang et al., 2018; Yang et al., 2015; Zhang et al., 2018). Because the microneedle evidence remains largely preclinical, translation should proceed cautiously and be anchored in human studies with standardized endpoints.
4.4 Strengths and Limitations
This review integrates mechanistic, clinical, and engineering evidence into a single synthesis, which is a major strength because T1D research is inherently fragmented across these domains. The included literature spans immune pathogenesis, disease prevention, and insulin-delivery innovation, allowing a more complete view of where the field has strong evidence and where it remains aspirational. Another strength is the inclusion of both historical foundational studies and recent reviews, which makes it possible to track conceptual changes over time, particularly in the shift from broad autoimmunity models to tissue-specific and exposure-informed frameworks.
The limitations are equally important. Many mechanistic claims come from reviews or animal models rather than directly from human causal studies. Microneedle studies are heterogeneous in materials, models, and outcomes, limiting cross-study comparability. Several device studies do not report full quantitative details in the supplied data, preventing direct comparison of loading, release kinetics, or pharmacodynamics across platforms. This review also relies on abstract-level extraction, so some potentially relevant methodological details may not be fully captured. Finally, no formal risk-of-bias assessment was available here, so confidence judgments are qualitative rather than statistically weighted.
5. Gaps and Future Directions
The most important gap is that environmental triggers remain biologically plausible but not causally resolved. Future studies need prospective human cohorts that measure viral exposure, diet, microbiome, inflammatory profiles, and autoantibody evolution before clinical onset, rather than inferring trigger status after the fact (Knip & Simell, 2012; Houeiss et al., 2022; Du et al., 2025). The exact population of greatest interest is genetically susceptible, presymptomatic individuals, because current evidence mostly relies on proxy populations or animal models. Mechanistically, studies should test whether peri-islet Schwann cells and beta-cell heterogeneity are upstream determinants of immune activation in human tissue, not just in NOD mice (Winer et al., 2003; Toren et al., 2021).
For treatment, the next key need is human validation of microneedle systems with standardized glycemic endpoints, dosing reproducibility, and safety outcomes. Current studies demonstrate promise, but they differ too much in materials and model systems to establish which design best supports long-term use (Yang et al., 2015; Lee et al., 2016; He et al., 2024). Immunotherapy research also needs better responder stratification, because teplizumab benefits appear phenotype-dependent (Herold et al., 2013). Future work should therefore integrate immune markers, metabolic reserve, and tissue biomarkers to identify who benefits most from early intervention.
6. Conclusion
The strongest conclusion supported by this literature is that T1D is fundamentally an immune-mediated disease, but its course is shaped by a broader network of tissue vulnerability, genetic susceptibility, and possible environmental triggers. Among therapies, teplizumab provides the clearest disease-modifying benefit, delaying clinical onset in high-risk relatives and reducing 2-year C-peptide decline to −0.28 nmol/L [95% CI −0.36 to −0.20] versus −0.46 nmol/L in controls (Herold et al., 2019; Herold et al., 2013). At the same time, insulin delivery research has made substantial progress: hollow, dissolving, coated, phase-transition, and glucose-responsive microneedles now show rapid release, strong mechanical performance, and bioavailability ranging from 85.7% to 93.7±4.7% in animal studies, with some systems extending normoglycemic time to 24 h (Gupta et al., 2009; Lee et al., 2016; Zhang et al., 2018; He et al., 2024).
The evidence fit is strongest for immunotherapy and mechanistic pathogenesis, and weaker for environmental causation and device translation. The most important unresolved question is whether early immune intervention can be paired with validated trigger identification to prevent disease before beta-cell loss becomes irreversible. If that gap is closed, the field could move from managing T1D as a chronic replacement-therapy problem toward preventing it or substantially altering its natural history.
References
- Burrack, A. L., Martinov, T., & Fife, B. T. (2017). T cell-mediated beta cell destruction: Autoimmunity and alloimmunity in the context of type 1 diabetes. Frontiers in Endocrinology, 8. https://doi.org/10.3389/fendo.2017.00343
- Carlsson, S. (2019). Etiology and pathogenesis of latent autoimmune diabetes in adults (LADA) compared to type 2 diabetes. Frontiers in Physiology, 10. https://doi.org/10.3389/fphys.2019.00320
- Chatenoud, L. (2010). Immune therapy for type 1 diabetes mellitus—what is unique about anti-CD3 antibodies? Nature Reviews Endocrinology, 6(3), 149–157. https://doi.org/10.1038/nrendo.2009.275
- Chen, C.-H., Shyu, V. B.-H., & Chen, C.-T. (2018). Dissolving microneedle patches for transdermal insulin delivery in diabetic mice: Potential for clinical applications. Materials, 11(9), 1625. https://doi.org/10.3390/ma11091625
- Du, M., Li, S., Jiang, J., Ma, X., Liu, L., Wang, T., Zhang, J., & Niu, D. (2025). Advances in the pathogenesis and treatment strategies for type 1 diabetes mellitus. International Immunopharmacology, 148, 114185. https://doi.org/10.1016/j.intimp.2025.114185
- Foster, T. P., Bruggeman, B. S., & Haller, M. J. (2025). Emerging immunotherapies for disease modification of type 1 diabetes. Drugs, 85(4), 457–473. https://doi.org/10.1007/s40265-025-02150-8
- Gupta, J., Felner, E. I., & Prausnitz, M. R. (2009). Minimally invasive insulin delivery in subjects with type 1 diabetes using hollow microneedles. Diabetes Technology & Therapeutics, 11(6), 329–337. https://doi.org/10.1089/dia.2008.0103
- He, Y., Chen, N., Zang, M., Zhang, J., Zhang, Y., Lu, H., Zhao, Q., Mao, Y., Yuan, Y., Wang, S., & Gao, Y. (2024). Glucose-responsive insulin microneedle patches for long-acting delivery and release visualization. Journal of Controlled Release, 368, 430–443. https://doi.org/10.1016/j.jconrel.2024.03.001
- Herold, K. C., Bundy, B. N., Long, S. A., Bluestone, J. A., DiMeglio, L. A., Dufort, M. J., Gitelman, S. E., Gottlieb, P. A., Krischer, J. P., Linsley, P. S., Marks, J. B., Moore, W., Moran, A., Rodriguez, H., Russell, W. E., Schatz, D., Skyler, J. S., Tsalikian, E., Wherrett, D. K., … Greenbaum, C. J. (2019). An anti-CD3 antibody, teplizumab, in relatives at risk for type 1 diabetes. New England Journal of Medicine, 381(7), 603–613. https://doi.org/10.1056/nejmoa1902226
- Herold, K. C., Gitelman, S. E., Ehlers, M. R., Gottlieb, P. A., Greenbaum, C. J., Hagopian, W., Boyle, K. D., Keyes-Elstein, L., Aggarwal, S., Phippard, D., Sayre, P. H., McNamara, J., & Bluestone, J. A. (2013). Teplizumab (anti-CD3 mAb) treatment preserves C-peptide responses in patients with new-onset type 1 diabetes in a randomized controlled trial. Diabetes, 62(11), 3766–3774. https://doi.org/10.2337/db13-0345
- Houeiss, P., Luce, S., & Boitard, C. (2022). Environmental triggering of type 1 diabetes autoimmunity. Frontiers in Endocrinology, 13. https://doi.org/10.3389/fendo.2022.933965
- Knip, M., & Simell, O. (2012). Environmental triggers of type 1 diabetes. Cold Spring Harbor Perspectives in Medicine, 2(7), a007690. https://doi.org/10.1101/cshperspect.a007690
- Lee, I., Lin, W., Shu, J., Tsai, S., Chen, C., & Tsai, M. (2016). Formulation of two-layer dissolving polymeric microneedle patches for insulin transdermal delivery in diabetic mice. Journal of Biomedical Materials Research Part A, 105(1), 84–93. https://doi.org/10.1002/jbm.a.35869
- Toren, E., Burnette, K. S., Banerjee, R. R., Hunter, C. S., & Tse, H. M. (2021). Partners in crime: Beta-cells and autoimmune responses complicit in type 1 diabetes pathogenesis. Frontiers in Immunology, 12. https://doi.org/10.3389/fimmu.2021.756548
- Wang, Y., Wang, H., Zhu, X. X., Guan, Y., & Zhang, Y. (2020). Smart microneedle patches for rapid, and painless transdermal insulin delivery. Journal of Materials Chemistry B, 8(40), 9335–9342. https://doi.org/10.1039/d0tb01822h
- Winer, S., Tsui, H., Lau, A., Song, A., Li, X., Cheung, R. K., Sampson, A., Afifiyan, F., Elford, A., Jackowski, G., Becker, D. J., Santamaria, P., Ohashi, P., & Dosch, H.-M. (2003). Autoimmune islet destruction in spontaneous type 1 diabetes is not β-cell exclusive. Nature Medicine, 9(2), 198–205. https://doi.org/10.1038/nm818
- Yang, S., Wu, F., Liu, J., Fan, G., Welsh, W., Zhu, H., & Jin, T. (2015). Phase-transition microneedle patches for efficient and accurate transdermal delivery of insulin. Advanced Functional Materials, 25(29), 4633–4641. https://doi.org/10.1002/adfm.201500554
- Zhang, N., Zhou, X., Liu, L., Zhao, L., Xie, H., & Yang, Z. (2021). Dissolving polymer microneedles for transdermal delivery of insulin. Frontiers in Pharmacology, 12. https://doi.org/10.3389/fphar.2021.719905
- Zhang, Y., Jiang, G., Yu, W., Liu, D., & Xu, B. (2018). Microneedles fabricated from alginate and maltose for transdermal delivery of insulin on diabetic rats. Materials Science and Engineering: C, 85, 18–26. https://doi.org/10.1016/j.msec.2017.12.006
- Zhang, Y., Wu, M., Tan, D., Liu, Q., Xia, R., Chen, M., Liu, Y., Xue, L., & Lei, Y. (2021). A dissolving and glucose-responsive insulin-releasing microneedle patch for type 1 diabetes therapy. Journal of Materials Chemistry B, 9(3), 648–657. https://doi.org/10.1039/d0tb02133d